
{kind=link}
BREAKING: Double‑Strand Breaks Prompt De Novo Chromatin Loops in Human Cells
Table of Contents
- 1. BREAKING: Double‑Strand Breaks Prompt De Novo Chromatin Loops in Human Cells
- 2. Key Findings in Minutes
- 3. Customary Cohesin Loops vs. DSB‑Induced loops
- 4. Why This Matters
- 5. Evergreen Insights
- 6. Reader Interaction
- 7. Frequently Asked Questions
- 8. okay, here’s a breakdown of the provided text, focusing on key information adn potential question/answer pairings, suitable for study or quiz readiness.I’ll categorize it for clarity.
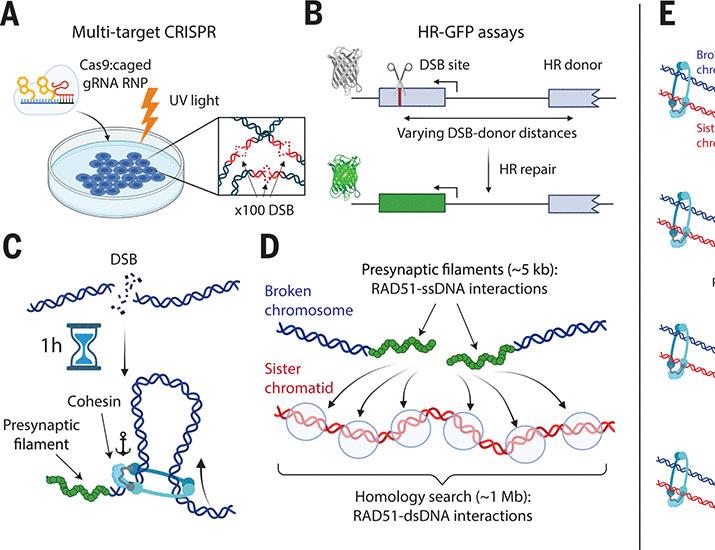
- 9. Cohesin Orchestrates Chromatin Scanning During RAD51‑Mediated Homology Search
- 10. RAD51‑Mediated Homology Search: Core Concepts
- 11. RAD51 Nucleoprotein Filament Formation
- 12. Key Steps in the Homology Search Process
- 13. Cohesin Complex – Architecture and Cellular Functions
- 14. Core Subunits of Cohesin
- 15. Cohesin Loading, Stabilization, and Release
- 16. Mechanistic Link Between Cohesin and RAD51 Homology Search
- 17. Cohesin‑Driven Chromatin Loop Extrusion
- 18. Spatial Proximity of Sister chromatids
- 19. Experimental evidence (2023‑2024)
- 20. Benefits of Cohesin‑Facilitated Scanning for DNA Repair
- 21. Practical Tips for Investigating Cohesin‑RAD51 Interactions
- 22. Case study: Cohesin Mutations, Cancer, and Homologous Recombination
- 23. Emerging Technologies for Visualizing Chromatin Scanning
– A team of molecular biologists has discovered that double‑strand breaks (DSBs) spark the rapid formation of fresh chromatin loops,positioning the loop base directly at the damage site.
Key Findings in Minutes
Researchers observed that when a DSB occurs, cohesin complexes swiftly anchor at the break, pulling neighboring DNA into a loop.
The newly formed loops differ from the static loops traditionally attributed to cohesin’s role in genome organization.
Customary Cohesin Loops vs. DSB‑Induced loops
| Feature | Traditional Cohesin Loops | DSB‑Induced De Novo Loops |
|---|---|---|
| Formation Trigger | Programmed genome folding during interphase | Spontaneous DNA double‑strand breaks |
| Loop Base Position | Pre‑defined CTCF sites | Exact DSB location |
| Stability | Relatively stable throughout cell cycle | Transient, resolves as repair completes |
| Biological role | Regulate transcription, replication timing | Facilitate homology‑directed repair |
Why This Matters
The discovery links genome architecture directly to the DNA damage response, suggesting that loop formation is a rapid, adaptive strategy to bring repair factors into proximity.
Understanding this mechanism could improve genome‑editing precision and inform cancer‑therapy approaches that exploit DSB repair pathways.
Evergreen Insights
Chromatin looping is orchestrated by cohesin’s ATP‑driven extrusion activity, a process first described in 2017 and continuously refined with high‑resolution Hi‑C data (Nature).
Recent 2024 studies show that loop extrusion speed can be modulated by post‑translational modifications of cohesin subunits, adding another layer of regulatory complexity (Cell).
These insights underscore the dynamic nature of the genome, where structural changes can be both cause and outcome of cellular events.
Reader Interaction
What implications do you think DSB‑induced loops have for gene therapy safety?
Have you observed unexpected DNA repair patterns that might be explained by transient looping?